
Eficiencia de la red en el trastorno del espectro autista y su relación con el crecimiento excesivo del cerebro
 John D. Lewis 1 * , Rebecca J. Theilmann 2 , Jeanne Townsend 3,4 y Alan C. Evans 1
John D. Lewis 1 * , Rebecca J. Theilmann 2 , Jeanne Townsend 3,4 y Alan C. Evans 1- 1 McConnell Cerebro Imaging Center, Montreal Neurological Institute, de la Universidad McGill, Montreal, QC, Canadá
- 2 Departamento de Radiología de la Universidad de California, San Diego, La Jolla, CA, EE.UU.
- 3 Departamento de Neurociencia de la Universidad de California, San Diego, La Jolla, CA, EE.UU.
- 4 Investigación sobre el Envejecimiento y el Desarrollo Laboratory de la Universidad de California, San Diego, La Jolla, CA, EE.UU.
Un importante cuerpo de evidencia vincula las diferencias en el tamaño del cerebro a las diferencias en la organización cerebral. Tenemos la hipótesis de que el aspecto del desarrollo de esta relación juega un papel en el trastorno del espectro autista (TEA), un trastorno del desarrollo neurológico que implica alteraciones en el crecimiento del cerebro. Los niños con TEA tienen cerebros anormalmente grandes por el segundo año de vida, y durante varios años a partir de entonces su tamaño cerebral puede haber múltiples desviaciones estándar por encima de la norma. Los mayores retrasos en la conducción y los costos de celulares presumiblemente asociados con las largas conexiones de larga distancia en estos cerebros más grandes se cree que influir en los procesos de desarrollo, dando lugar a una organización cerebral alterada con menos comunicación entre regiones espacialmente distantes. Esto ha sido apoyado por modelos computacionales y por los hallazgos que vinculan mayor volumen intracraneal, un índice de máxima-el tamaño del cerebro durante el desarrollo, a la reducción de la conectividad interhemisférica en individuos con TEA.En este trabajo, evaluamos esta hipótesis a través de un análisis de todo el cerebro de la eficiencia de la red. Utilizamos tractography difusión para estimar la fuerza y la longitud de las conexiones entre todos los pares de regiones corticales. Calculamos la eficiencia de la comunicación entre todos los nodos de la red y todos los demás, y dentro de los vecindarios locales, que luego evaluamos la relación de estas medidas con el volumen intracraneal, y las diferencias en estas medidas entre los adultos con autismo y controles típicos. Volumen intracraneal se demuestra que es inversamente proporcional a la eficiencia de las regiones extendidas de la corteza. Por otra parte, los patrones espaciales de las reducciones en la eficiencia en el autismo tienen un parecido sorprendente a las relaciones regionales entre la eficiencia y el volumen intracraneal, sobre todo por la eficiencia local. Así, los resultados proporcionan apoyo adicional a la hipótesis de vínculo entre el crecimiento excesivo del cerebro en niños con autismo y la eficiencia de la organización del cerebro en los adultos con autismo.
Introducción
Los cerebros difieren dramáticamente en tamaño y estructura a través de especies. Estas dos dimensiones de la variación no son independientes, sino grandes cerebros no son grandes cerebros pequeños. La organización de ambos-sustancia blanca-gris y varía con el tamaño del cerebro, pero no de una manera uniforme. Aumentar el tamaño del cerebro está asociada con un mayor de la materia blanca y gris-materia ratio ( Rilling y Insel, 1999b ; Zhang y Sejnowski, 2000 ), pero un grado reducido de conectividad de larga distancia ( Ringo, 1991 ; Rilling y Insel, 1999a ; Karbowski, 2003 ; Changizi, 2007 ), así como con el aumento de estructura modular ( Changizi y Shimojo, 2005 ), mayor convolutedness superficie ( Jerison, 1982 ; Prothero y Sundsten, 1984 ; Hofman, 1985 ), y varios otros aspectos morfológicos y celulares de la organización neuronal. Leyes de escala capturan gran parte de la variación en la estructura en términos de tamaño del cerebro ( Jerison, 1982 ; Ringo, 1991 ; Karbowski, 2003 ; Changizi y Shimojo, 2005 ; Changizi, 2007 ). Sin embargo, la significativa variabilidad estructural permanece en paradero desconocido por las leyes de escala.
Las bases de estas relaciones de escala no se conocen bien, pero se cree que están relacionados con un principio de diseño originalmente postulada por Ramón y Cajal: que el diseño de circuitos neuronales se encuentra bajo presión para reducir al mínimo los costos de celulares y retrasos de la conducción ( Ramón y Cajal, 1995 ). El aumento de tamaño del cerebro proporciona una mayor potencia de cálculo, pero al enorme aumento de los costes. Material de neural es caro de construir y de operar. El cerebro humano constituye sólo aproximadamente el 2 por ciento del peso total del cuerpo, pero su funcionamiento es responsable de aproximadamente el 15 por ciento del gasto cardíaco, 20 por ciento del uso de oxígeno, y 25 por ciento del uso de glucosa ( Magistretti, 1999 ). Estos costos metabólicos son en gran parte debido al coste de la señalización neuronal, y el mantenimiento de los potenciales de reposo necesarios para la señalización neuronal. Estos costes aumentan con la zona de superficie de la membrana, que aumenta con el número y tamaño de los axones. Cerebros más grandes tienen un mayor número de axones, y el más largo de estos axones son tanto más largo y ligeramente más grande en diámetro que son los de cerebros más pequeños, por lo que el área total de superficie de la membrana se incrementa. Diámetro del axón no aumenta suficientemente con el tamaño del cerebro, sin embargo, para compensar el aumento de las longitudes de las fibras, de modo cerebros más grandes también tienen retrasos de la conducción más largos ( Olivares et al., 2001 ). Estos mayores costos y retrasos en la conducción parecen estar relacionados con, al menos, algunos de los aspectos de la organización que se escalan con el tamaño del cerebro, por ejemplo, la reducción del grado de conectividad a larga distancia ( Ringo, 1991 ; Rilling y Insel, 1999a ; Karbowski, 2003 ; Changizi , 2007 ).
El enfoque en las diferencias entre especies, donde las diferencias en el tamaño del cerebro puede contener más de 1000 veces dentro de las clases, por ejemplo, Mammalia, y 100 veces en órdenes, por ejemplo, primates, permite relaciones entre el tamaño y la estructura del cerebro a ser aparente a pesar de las diferencias en la estructura no relacionada con el tamaño del cerebro. Pero, no tiene en cuenta las diferencias potencialmente importantes en las trayectorias de crecimiento cerebral del desarrollo. Existen diferencias entre especies importantes en la tasa de crecimiento del cerebro, y las trayectorias de desarrollo, incluso puede variar considerablemente entre los individuos, por ejemplo, el tamaño del cerebro puede diferir hasta en un 50% en los niños de la misma edad ( Giedd, 2008 ). Diferencias en el tamaño del cerebro entre los adultos representan algunas de las diferencias en la estructura ( Jancke et al, 1997. ; Miel et al, 2009. ; Lewis et al, 2009. ), las diferencias en las trayectorias de crecimiento cerebral probablemente expliquen la variabilidad estructural adicional.
Reorganización neuronal sustancial se produce durante el desarrollo. El desarrollo neuronal es en gran parte una combinación de exuberancia excesiva y eliminación basada en la competencia. Un gran número de proyecciones transitorios se producen durante el desarrollo cortical ( Rakic et al, 1986. ; LaMantia y Rakic, 1990 ) se conservan las conexiones, y que está determinada por sus demandas metabólicas y su capacidad para competir por las neurotrofinas ( Van Ooyen y Willshaw, 1999 ). Debido a la menor grado de mielinización en el cerebro en desarrollo que en el cerebro maduro, las diferencias en los retrasos de la conducción y los costos metabólicos asociados con diferencias en la longitud de la fibra serán sustancialmente mayor ( Chugani et al, 1987. ; Paus et al, 1999. ; Thatcher et al., 2008 ). Por lo tanto, en la medida en que las diferencias en el tamaño del cerebro durante el desarrollo coinciden con las diferencias en el tamaño del cerebro en individuos maduros, los procesos de desarrollo normales pueden ser la base de al menos una parte de las relaciones de escala visto a través y dentro de las especies, por otra parte, las diferencias en el tamaño del cerebro durante el desarrollo que no coinciden con las diferencias en el tamaño del cerebro en individuos maduros puede dar cuenta de una parte de la variabilidad estructural que no se explica por la ampliación leyes.
Esta conjetura es claramente relevante para los trastornos del desarrollo que muestran anomalías en las trayectorias de crecimiento del cerebro. Trastorno del espectro autista (ASD) es un caso así. TEA es un trastorno del desarrollo neural definida por alteraciones en las interacciones sociales recíprocas, deficiencias en la comunicación verbal y no verbal, y un repertorio restringido de actividades e intereses (American Psychiatric Association, 1994). La etiología de la TEA es desconocida, pero ahora hay consenso en que el tamaño del cerebro durante el desarrollo se incrementa. Los bebés que van a un diagnóstico de TEA muestran el crecimiento del cerebro anormalmente rápido durante los primeros años de vida ( Lainhart et al, 1997. ; Redcay y Courchesne, 2005 ), y después del segundo o tercer año de vida los niños con TEA muestran una mayor cabeza tamaño ( Lainhart et al, 1997. ; . Hazlett et al, 2005 ) y el tamaño del cerebro ( Piven et al, 1995. ; Courchesne et al, 2001. ; . Hazlett et al, 2005 ). Temprano en el desarrollo de esta diferencia de tamaño puede haber múltiples desviaciones estándar por encima de la norma ( Redcay y Courchesne, 2005 ).
Lewis y Elman (2008) han demostrado a través de modelos computacionales que el aumento de retrasos de la conducción presumiblemente asociados con el crecimiento excesivo del cerebro temprano en ASD pueden llevar a la underconnectivity largo alcance funcional y estructural posterior. Además, en los adultos con TEA, Lewis et al. (2012) han demostrado que la longitud del cuerpo calloso tracto ajustado para el volumen intracraneal (ICV), un índice de máximo-tamaño del cerebro durante el desarrollo ( Whitwell et al, 2001. ; . Aylward et al, 2002 ; . Buckner et al, 2004 ) , muestra la relación inversa típica relativa al tamaño de cuerpo calloso, y por lo que el crecimiento excesivo temprano del cerebro en el autismo parece en cuenta hecho por alguna porción de la tarde underconnectivity largo rango observado.
En el presente trabajo hemos ampliado este trabajo para evaluar el impacto del crecimiento excesivo del cerebro temprano en los TEA en la organización general del cerebro. Se realizó un análisis de la red y se evaluó la relación entre los parámetros de red y ICV. Métodos de análisis de la red han evolucionado en la última década y media, desde aplicaciones sencillas de la teoría de grafos, que evalúan sólo la topología de red ( Watts y Strogatz, 1998 ), con los enfoques más sofisticados que tienen en cuenta los aspectos espaciales de la conectividad para evaluar la eficiencia de transferencia de información dentro de la red ( Latora y Marchiori, 2001 , 2003 ; Achard y Bullmore, 2007 ; Bullmore y Sporns, 2012 ). Estos enfoques utilizan medidas de la longitud y la fuerza de las conexiones entre todos los pares de regiones anatómicas para estimar la eficiencia con la información puede transferirse entre regiones. Se utilizó tractography probabilístico para estimar la fuerza de la conectividad entre todos los pares de regiones, y la longitud de las conexiones entre las regiones. Hemos calculado la eficiencia de la comunicación de todas las regiones a todos los otros, y dentro de los vecindarios locales. Luego se evaluó la relación entre estas dos medidas de eficiencia y de ICV, así como las diferencias de grupo en la eficiencia. Nosotros predijimos que habría una relación inversa entre ICV y las dos medidas de eficiencia, lo que refleja un efecto adverso del crecimiento excesivo del cerebro en la organización general del cerebro, y que esto explicaría una parte de las diferencias entre los grupos en la eficiencia.
Métodos
Los participantes
Un total de 44 hombres adultos participaron en el estudio: 22 con TEA que oscila entre 19 y 51 años de edad (media 34,14; SD 10.67) y 22 varones adultos típicos oscilan entre 20 y 45 años de edad (media 32,25; SD 9.98) . Todos los participantes se reunieron ASD criterios diagnósticos para ASD en el DSM-IV según lo confirmado por un médico licenciado. Dieciocho de los veintidós participantes ASD se reunió con el diagnóstico DSM para el trastorno autista (autismo clásico) y, basándose en la ausencia de retraso en el lenguaje temprano y ninguna anomalía significativa en la comunicación, cuatro de los veinte y dos sujetos se reunieron, además, los criterios diagnósticos para el trastorno de Asperger. Autism Diagnostic Interview, revisado puntuaciones (ADI-R) estaban disponibles para 16 de los participantes ASD; puntajes Autism Diagnostic Calendario Observación (ADOS) estaban disponibles para 18, y puntuaciones de Autismo Infantil Scale (CARS) estaban disponibles para 12. Tabla 1 se resumen estos datos. En todos menos uno de los casos el diagnóstico de TEA fue confirmada por todas las evaluaciones adicionales disponibles, la única excepción fue por debajo del punto de corte para los coches, pero por encima de todos los puntos de corte para el ADI-R y ADOS. La capacidad intelectual general de los participantes ASD fue evaluada por la Escala Revisada de Inteligencia de Wechsler para Adultos (WAIS-R) o la Escala Wechsler de Inteligencia Abreviada (WASI). Las puntuaciones medias fueron: CI verbal, 88,48 ± 23,06; IQ Performance, 106,10 ± 15,91. Las personas con antecedentes de trastornos médicos o neurológicos significativos, como convulsiones o con el síndrome de X frágil se excluyeron de la muestra. Participantes típicos con un pariente de primer grado con un diagnóstico de TEA fueron excluidos de la muestra. Los participantes fueron los de Lewis et al. (2012) aumentada por nuevos datos de las personas con TEA. Aquellos sujetos que eran capaces dieron su consentimiento informado, un cuidador dio su consentimiento informado para los demás. El estudio fue aprobado por el Programa de Protección de la Investigación Humanos de la Universidad de California, San Diego.
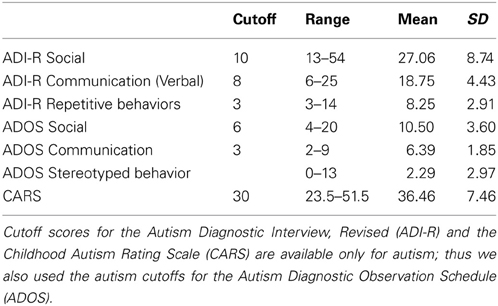
Tabla 1. Los datos sobre el comportamiento .
Imaging
Todos los sujetos fueron escaneados en el Centro de UCSD para fMRI en un GE Signa EXCITE 3.0T escáner de corto calibre con una bobina de cabeza arreglo de ocho canales. Hay tres tipos de imágenes fueron adquiridas de cada sujeto: (i) un conjunto de 3D T 1 imágenes ponderadas (Fast Gradient Echo, SPGR; TE = 3,1 ms; flip = ángulo 12; NEX = 1; FOV = 25 cm; matriz = 256 × 256), (ii) dos conjuntos de imágenes ponderadas de difusión isotrópica distribuidos a lo largo de 15 direcciones (doble spin-eco, EPI; TR = 15 s; TE = 89 ms; 45 cortes axiales; NEX = 2; FOV = 22 cm; matriz = 128 × 128; resolución = 1,875 × 1,875 × 3 mm; 3 mm intercalados cortes contiguos; b valor = 1400 s / mm 2 ), y (iii) fieldmaps adaptadas a las imágenes de difusión ponderada. Durante las exploraciones de adquisición se inspeccionaron visualmente para asegurarse de que se recogieron datos utilizables. Cuando el movimiento introdujo artefactos visibles en varios volúmenes, la secuencia de escaneo se ha cancelado y reiniciado, o una exploración adicional fue adquirida. Tenga en cuenta que al menos dos conjuntos de imágenes ponderadas de difusión fueron adquiridas, cada uno con NEX = 2, por lo que cada imagen se adquirió al menos cuatro veces. Fieldmaps fueron adquiridos antes que se adquirieron las primeras imágenes potenciadas en difusión, y, en los casos en que había entre el movimiento de exploración, un conjunto adicional de fieldmaps fue adquirida después de la segunda.
Procesamiento de imágenes
Los T 1 volúmenes fueron procesados con CIVET, una tubería de análisis de imagen estructural totalmente automatizado desarrollado en el Instituto Neurológico de Montreal. CIVET corrige intensidad no uniformidades utilizando N3 ( trineo et al, 1998. ); alinea los volúmenes de entrada a la plantilla de ICBM-152-NL-Talairach como ( Collins et al, 1994. ); clasifica la imagen en la materia blanca, la materia gris , líquido cefalorraquídeo, y el fondo ( Zijdenbos et al, 2002. ; . Tohka et al, 2004 ), y extrae la materia blanca y las superficies pial ( Kim et al, 2005. ). ICV se calculó a través de los atlas basados procedimiento de normalización espacial describe en Buckner et al. (2004) . Los resultados Civet se inspeccionaron visualmente para asegurarse de que la construcción de la superficie era correcta, y luego usaron para construir la semilla, detener y máscaras de destino para su uso con la FSL probtrackx ( Behrens et al., 2007 ). Máscaras de semillas de control desde el cual voxels extensiones se siembran; máscaras semillas eran de la materia blanca. Deje de máscaras determinan dónde se detiene la propagación del tracto; máscaras de parada fueron los voxels en el límite de la materia blanca. Máscaras objetivo determinar la asignación de los voxels de las máscaras escalas a regiones del cerebro; máscaras objetivo eran los voxels en el límite de la materia blanca y la corteza, y se asignan estos voxels al Anatómico Etiquetado automático (AAL) atlas ( Tzourio-Mazoyer et al ., 2002 ), que se muestra en la Figura 1 .
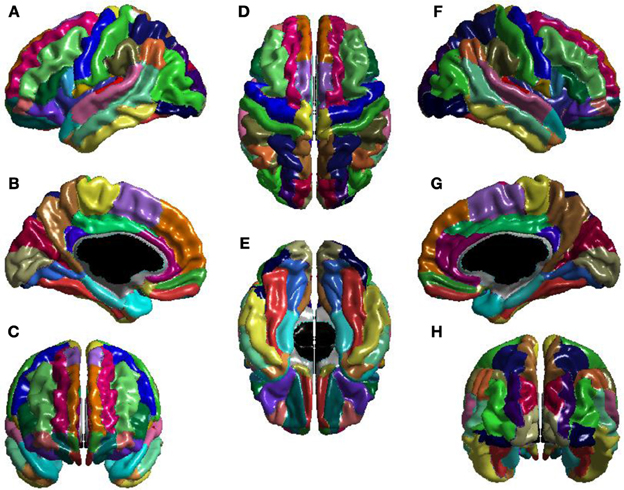
Figura 1. Los AAL atlas ha sido visto: (A) a la izquierda lateral, (B) dejaron medial, (C) anterior, (D) superior, (E) inferior, (F) lateral derecha, (G) medial derecho, y (H) posterior. Las regiones son de color para identificar los límites de la región. Similitud de colores entre las regiones separadas espacialmente no es significativo; todas las regiones están espacialmente contiguas. La parcelación cortical se basa en las referencias anatómicas, por ejemplo, los surcos. Hay 78 regiones corticales. Estas regiones corticales sirven como los nodos de la red.
Cada conjunto de datos de difusión se corrigió por primera vez de las distorsiones causadas por la falta de homogeneidad en el campo magnético utilizando los fieldmaps. Esto se hizo utilizando el software desarrollado por el Centro de UCSD para fMRI. Los volúmenes de difusión ponderada resultantes se someten a continuación a una evaluación cuantitativa de control de calidad usando DTIprep ( Liu et al., 2010 ). DTIprep corrige los artefactos de movimiento cuando sea posible, y excluye las direcciones de los datos cuando la corrección no es posible. Para cada tema, se eligieron los dos volúmenes en difusión con el menor número de direcciones excluidas para su posterior procesamiento. Los b 0 volúmenes de ambas exploraciones de difusión fueron entonces afín registrado a la T 1 -volumen en el espacio estereotáxico usando el Oxford University FMRIB software de Biblioteca (FSL) ligar ( Jenkinson y Smith, 2001 ), y la resultante transforma utiliza para alinear los dos 4D volúmenes; el componente de rotación se aplicó a los vectores direccionales. Los dos eran entonces fusionado usando de FSL fslmerge . A continuación, el volumen combinado que se procesan para tractography probabilístico con la FSL bedpostx ( Behrens et al., 2007 ). Tractography Probabilístico, utilizando de FSL probtrackx con corrección de distancia-bias ( Behrens et al., 2003 , 2007 ), y luego se sembró de 10.000 lugares al azar dentro de cada voxel de las máscaras de semillas para generar el número de tramos que conectan voxels en la máscara destino. Un volumen de difusión 4D escala nativa se generó usando el mismo procedimiento, pero con el componente de escala retirado de las transformadas; este se procesó de la misma manera para generar las longitudes de las conexiones entre elementos de imagen volumétrica en la máscara destino. Estos resultados fueron luego compilados para cada región AAL matrices generadoras del número total de conexiones entre cada par de regiones de AAL, y la duración media de esas conexiones. El número total de conexiones entre cada par de regiones de AAL a continuación, se divide por el tamaño medio de las dos regiones AAL para proporcionar un índice de la fuerza de la conectividad entre pares de regiones.
Análisis
Se calculó la eficiencia de la comunicación para todas las regiones, con base en la definición establecida en Latora y Marchiori (2001 , 2003) . La relación de ICV a la eficiencia se evaluó con modelos lineales estadísticos, así como las diferencias de grupo en la eficiencia. Corrección para comparaciones múltiples se realizó mediante el método de la tasa de falso descubrimiento ( Benjamini y Hochberg, 1995 ).
Latora y Marchiori (2001) definen la eficiencia ε ij en la comunicación entre los nodos i y j a ser inversamente proporcional a la longitud más corta trayectoria d ij entre los nodos i y j . Toman la más corta longitud de la trayectoria d ij ser la suma más pequeña de las distancias físicas a través de todos los caminos posibles de i a j en el gráfico, es decir, la distancia de recorrido, no el número de bordes ni la distancia euclidiana. La eficiencia de una red, T , es luego
donde N es el número de nodos en el gráfico de la red T ; ε ij es la eficiencia de la conexión entre los nodos i y j , y d ij es la longitud de la ruta más corta en términos de distancias físicas, entre los nodos i y j . Esta medida se normaliza E ( T IDEAL ), la red totalmente conectada. Tenga en cuenta que las medidas de eficiencia tienen en cuenta las distancias físicas que intervienen en la transferencia de información, y por lo tanto se relacionan más estrechamente con los sustratos neurobiológicos de hacer medidas puramente topológicos ( Watts y Strogatz, 1998 ; Achard y Bullmore, 2007 ; Rubinov y Sporns, 2010 ) .
Latora y Marchiori (2001) aplican esta formulación tanto de toda la red, lo que ellos denominan como la eficiencia mundial , y para las subredes de los vecinos inmediatos de cada nodo, sino que definen la eficiencia local, como la media de E ( T i ), para todos los nodos i , donde T i es el subgrafo de todos los vecinos del nodo i . Estas definiciones dan una sola medida de la eficiencia local de y de la eficiencia mundial para toda la red. Pero, las definiciones se pueden dar traducciones directas para proporcionar medidas de eficiencia para cada nodo, o para las colecciones de nodos. Achard y Bullmore (2007) definen la eficiencia nodal , lo que nos referiremos como la eficiencia nodal mundial , como la inversa de la armónica media del número mínimo de bordes entre un nodo, i , y todos los otros nodos de la red. Utilizando las distancias físicas, como por Latora y Marchiori (2001) , la eficiencia global de ganglionar de nodo i es por lo tanto
donde N es el número de nodos en el gráfico de la red T ; ε ij es la eficiencia de la conexión entre los nodos i y j , y d ij es la longitud de la ruta más corta, en términos de distancias físicas, entre los nodos i y j . La definición de la eficiencia local de mismo modo se puede analizar para proporcionar una medida de la eficiencia local de ganglionar ; recordar que la eficiencia local de de una red es la media de E ( T i ), para todos los nodos i , donde T i es el subgrafo de todos los vecinos del nodo i . Por lo tanto, la eficiencia local de ganglionar de nodo i es simplemente
donde N Gi es el número de nodos en el subgrafo T i que consta de todos los vecinos de i; ε jk es la eficiencia de la conexión entre los nodos j y k , y d jk es la longitud del camino más corto, en términos de distancias físicas, entre los nodos j y k .
Estas definiciones tratan conexiones de una manera binarizada, es decir, ya sea como existente o no. Pero, los puntos fuertes de las conexiones reflejan, aunque pobremente, propiedades biofísicas de los axones subyacentes que están relacionados con la velocidad de conducción y los costos metabólicos, por ejemplo, la mielinización. Por otra parte, las conexiones débiles de largo alcance entre los módulos conectados fuertemente han argumentado para proporcionar los atajos que hacen que el cerebro de una eficiente arquitectura (small-world Gallos et al., 2012 ). Los puntos fuertes de las conexiones en el cerebro pueden así ser fundamental para una evaluación exacta de su eficacia. Por lo tanto, utilizamos una versión de estas medidas que incorpora el peso de conexión, es decir, el número total de vías que conectan dos regiones, corregidos para el tamaño de la distancia-el sesgo y la región. Basado en Rubinov y Sporns (2010) , se define el la distancia ponderada entre los nodos i y j como
donde S es el conjunto de bordes en el camino más corto entre los nodos i y j; l e es la longitud de borde e , y W e es el peso de conexión para el borde de correo . También basado en Rubinov y Sporns (2010) , nuestras fórmulas ponderadas de la eficiencia global de nodal y eficiencia locales nodal son
donde N es el número de nodos en el gráfico de la red G; y d W ij es el camino más corto, en términos de distancia ponderada , entre los nodos i y j ; y
donde N Gi es el número de nodos en el subgrafo T i que consta de todos los vecinos de i; es el camino más corto, en términos de distancia ponderada , entre los nodos j y k , y W ij y W ik son los pesos de conexión entre nodos i y j y i y k , respectivamente. Según Latora y Marchiori (2001) , estas medidas se normalizan considerando la red totalmente conectada.
El impacto de tamaño máximo cerebro durante el desarrollo en la eficiencia se evaluó, así como las diferencias de grupo en la eficiencia. Como por Lewis et al. (2012) , se utilizó ICV como un índice de tamaño máximo cerebro durante el desarrollo ( Whitwell et al, 2001. ; Aylward et al, 2002. ; Buckner et al, 2004. ). La relación entre ICV y la eficacia se evaluó a través de modelos lineales estadísticos, controlar la edad y el volumen total del cerebro. Las diferencias de grupo en la eficiencia fueron evaluados a través de modelos lineales estadísticos, controlar la edad. Diferencias entre los grupos potenciales en las relaciones entre ICV y las medidas de eficiencia se evaluaron considerando el término interacción grupo x ICV en modelos con ambos términos. En todos los casos, la corrección para comparaciones múltiples se realizó mediante el método de la tasa de falso descubrimiento ( Benjamini y Hochberg, 1995 ).
Resultados
La relación entre ICV y la eficiencia local de ganglionar se muestra en la Figura 2 . El t -estadística es negativo en toda la corteza, por lo tanto para todas las regiones se trata de una relación inversa: mayor ICV se asoció con una menor eficiencia locales nodal . Esta relación inversa es significativo sobre casi la totalidad de la parte posterior del cerebro, y también el lóbulo frontal del hemisferio derecho. La relación es notablemente menos negativa durante dorsal izquierda corteza frontal lateral, y no alcanzó significación sobre gran parte de la izquierda dorsal hemisferio cortical lateral, la relación inversa es más fuerte sobre la superficie medial, y es significativo en la mayor parte de la superficie medial de ambos hemisferios.
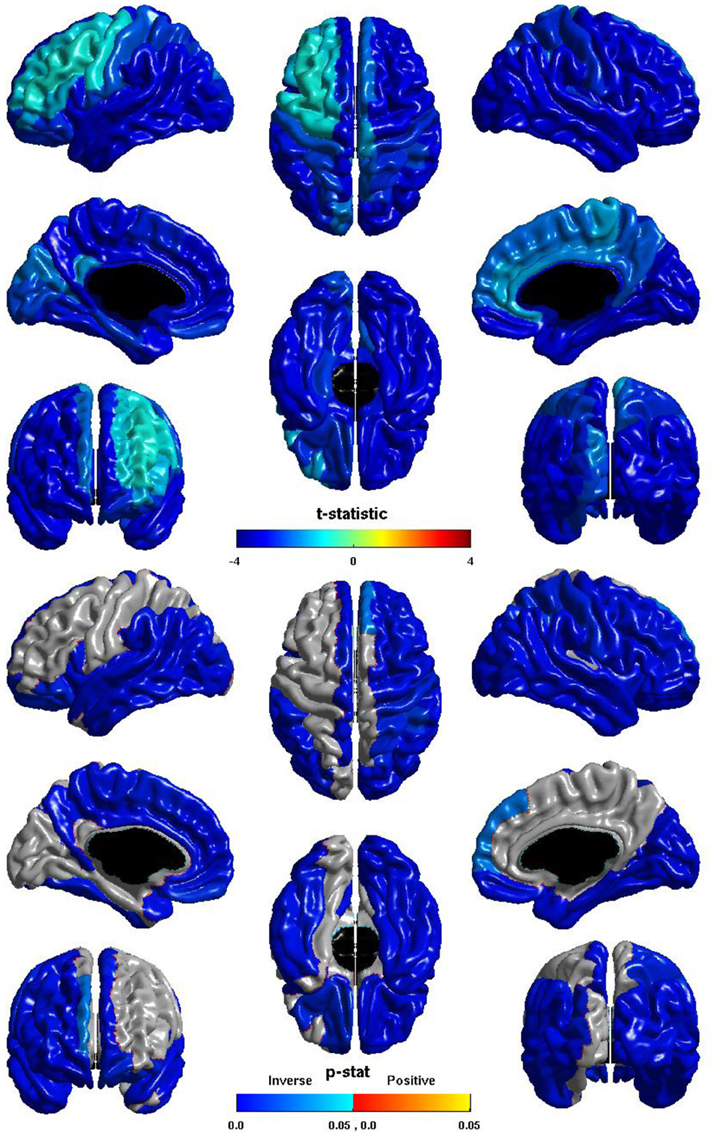
Figura 2. Eficiencia e ICV Nodal Local. El t -estadística (arriba) y el p -estadístico (abajo) para la relación entre ICV y eficiencia locales nodal en cada región del atlas AAL. Una negativa t -estadística representa la disminución de la eficacia local de nodal con el aumento de ICV. El t -estadística es abrumadoramente negativa. El p -estadística es corregida-FDR, y es de color azul, donde la relación inversa es significativo, y naranja en donde una relación positiva significativa. No hay regiones muestran una relación positiva significativa. Relaciones inversas significativas se observan de forma bilateral en los lóbulos temporales, las circunvoluciones angulares y supramarginal, los opercularis pars, la corteza frontal orbital y la circunvolución frontal superior, el hemisferio derecho muestra esta relación inversa más ampliamente en el frontal y el lóbulo parietal, el hemisferio izquierdo muestra la relación más extensa en la superficie medial.
Las diferencias de grupo en la eficiencia local de ganglionar se muestran en la Figura 3 . El t -estadística es negativo en toda la corteza, por lo tanto para todas las regiones eficiencia nodal locales se reduce en las personas con TEA. Esta reducción es significativa sobre casi la totalidad de la parte posterior del cerebro, y también el lóbulo frontal del hemisferio derecho. El t -estadística es visiblemente menos negativo sobre la corteza frontal lateral izquierda, y la diferencia entre los grupos no alcanzó significación en la mayor parte de la corteza frontal lateral izquierda, la diferencia es significativa con respecto a la mayor parte de la superficie medial izquierdo. La diferencia entre los grupos no es significativa para la mayor parte de la superficie anterior medial derecho a la cuneus. Tenga en cuenta que el patrón de diferencias de grupo en nodales eficiencia locales paralelos el de la relación inversa entre ICV y eficiencia locales nodal . La similitud del coseno de los dos t vectores-estadístico es 0.9848.
Figura 3. Eficiencia Nodal Local y Grupo. El t -estadística (arriba) y el p -estadístico (abajo) para el grupo de diferencia en la eficiencia local, nodal en cada región del atlas AAL. Una negativa t -estadística representa la eficiencia reducida en ASD. El t -estadística es negativa en todas partes. El p -estadística es corregida-FDR, y es de color azul, donde hay una reducción significativa en la eficacia local de nodal en ASD, y naranja donde hay un aumento significativo de la ASD. No hay regiones muestran aumentado significativamente la eficiencia local, nodal en ASD. Reducciones significativas son vistos de forma bilateral en lo temporal, occipital y lóbulos parietales, y en las opercularis pars, el hemisferio derecho se muestra, además, las reducciones sobre las regiones laterales del lóbulo frontal, el hemisferio izquierdo muestra más extensas reducciones sobre la superficie medial. Tenga en cuenta las similitudes con la relación de eficiencia locales nodal e ICV.
La relación entre ICV y la eficiencia global de ganglionar se muestra en la Figura 4 . El t -estadística es negativo durante la mayor parte de la corteza, por lo que este es de nuevo generalmente una relación inversa: mayor ICV se asoció con una menor eficiencia mundial nodal . Relaciones inversas significativas se observan en el hemisferio izquierdo en todos los lóbulos, sobre todo en la corteza visual, la pre-y post-central de circunvoluciones, y en la corteza auditiva primaria, las relaciones inversas significativas se observan en el hemisferio derecho en el lóbulo temporal, el precuneus, y el lóbulo paracentral, y las relaciones inversas significativas se observan de forma bilateral en la corteza cingulada y la corteza orbitofrontal.
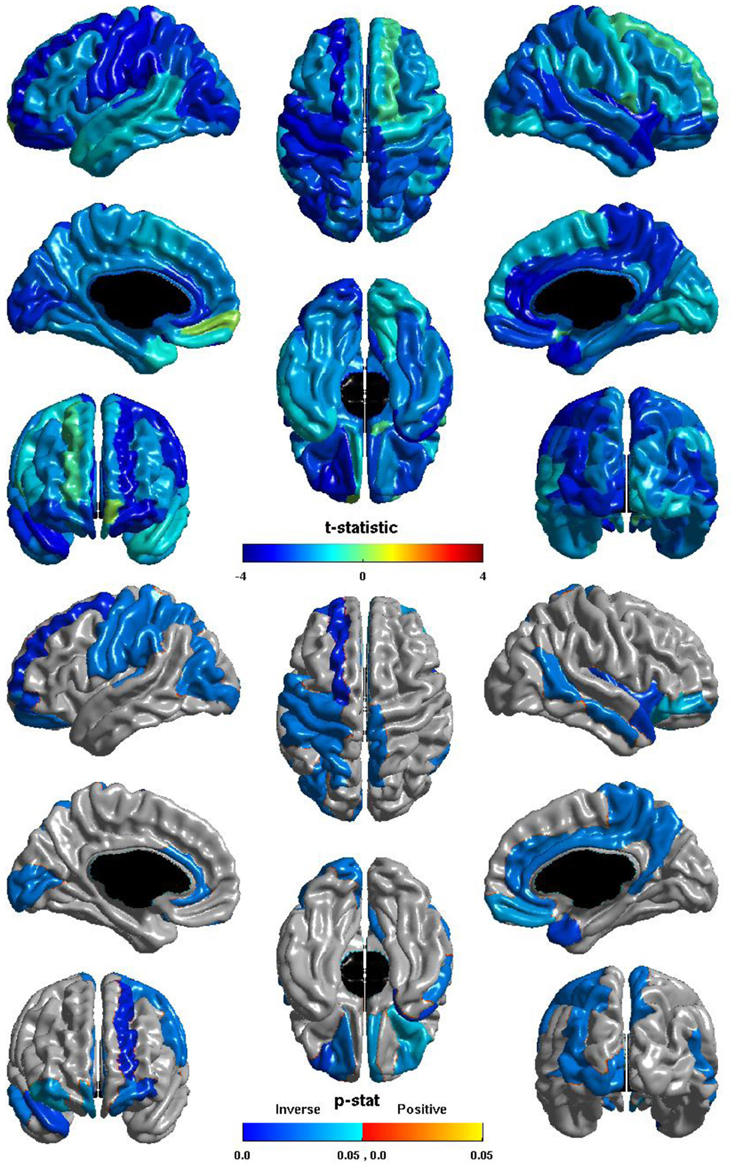
Figura 4. Eficiencia e ICV Nodal Global. El t -estadística (arriba) y el p -estadístico (abajo) para la relación entre ICV y la eficiencia global de nodal en cada región del atlas AAL. Una negativa t -estadística representa la disminución de la eficiencia global de nodal con el aumento de ICV. El t -estadística es predominantemente negativa. El p -estadística es corregida-FDR, y es de color azul, donde la relación inversa es significativo y naranja donde una relación positiva significativa. No hay regiones muestran una relación positiva significativa. Relaciones inversas significativas se observan en el occipital izquierdo, parietal, y los lóbulos frontales, y en la corteza auditiva primaria, en el lóbulo temporal, precuneus y paracentral lóbulo derecho, y de forma bilateral en la corteza cingulada y la corteza orbitofrontal.
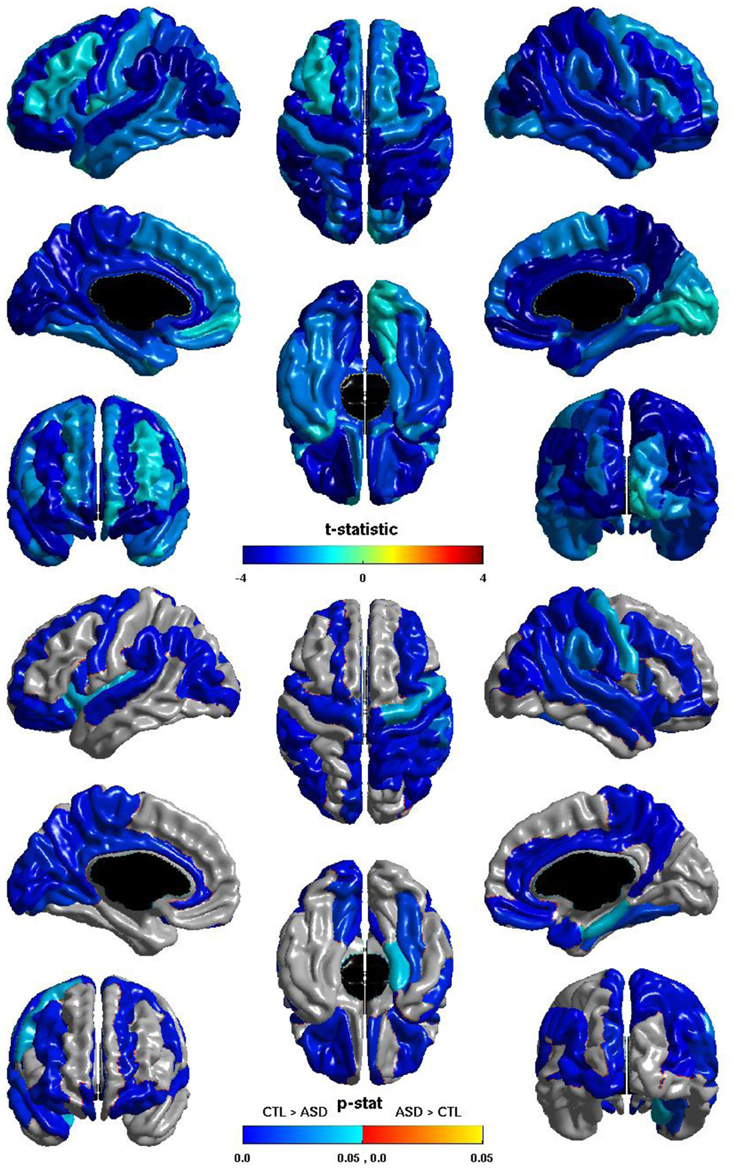
Las diferencias entre los grupos en la eficiencia global de ganglionar se muestran en la Figura 5 . El t -estadística es negativo en toda la corteza, por lo tanto para todas las regiones eficiencia nodal mundial se reduce en las personas con TEA. Esta reducción es significativa en las regiones de todos los lóbulos de ambos hemisferios. Tenga en cuenta que estas reducciones se superponen con los de la relación de ICV y la eficiencia global de ganglionar , pero son más extensa, especialmente en el hemisferio derecho. La similitud del coseno de los dos t vectores-estadístico es 0.9584.
Discusión
Las redes con un alto grado de conectividad espacial local, pero con pocas conexiones de largo alcance, es decir, los accesos directos, tienen una alta eficiencia local y la baja eficiencia global; redes con un alto grado de conectividad de largo alcance, pero que carecen de agrupamiento espacial local tienen alta eficiencia global y la baja eficiencia local. Los sistemas biológicos en las redes generales, y los nervios, en particular, tienden a equilibrar la eficiencia global con eficiencia local tiene una fuerte agrupación local mezclado con suficiente conectividad de largo alcance para permitir una rápida comunicación entre los nodos remotos, los cuales han sido denominados propiedades de "mundo pequeño" ( Watts y Strogatz, 1998 ; Latora y Marchiori, 2001 , 2003 ). La relación inversa que se muestra aquí entre ICV y ambos locales nodal y la eficiencia global de nodal sugiere que la desviación en el crecimiento del cerebro trayectorias impactos tanto en la comunicación a larga distancia y la comunicación dentro de la vecindad, y los impactos tanto de manera similar. La ausencia de un grupo * Interacción ICV en ambos casos indica que lo mismo es cierto tanto en adultos típicos y adultos con TEA. Las reducciones en tanto locales ganglionar y la eficiencia global de ganglionar visto en individuos con ASD se alinean con esta relación inversa, en combinación con el crecimiento excesivo del cerebro que se produce en TEA, que sugieren que el crecimiento excesivo del cerebro puede explicar al menos parte de las reducciones en la eficiencia; y la similitud del patrón espacial de las reducciones en la eficiencia con el patrón de las relaciones entre ICV y la eficiencia apoya aún más esta conclusión.
Estos resultados complementan nuestros trabajos anteriores que muestran una relación inversa entre la longitud ICV-ajustado de fibras del cuerpo calloso y el grado de conectividad interhemisférica en ASD ( Lewis et al., 2012 ), y nuestro trabajo de modelado computacional que muestra que el crecimiento excesivo del cerebro temprano en ASD puede hacer que las reducciones posteriores en la conectividad de largo alcance ( Lewis y Elman, 2008 ). Estos estudios sugieren que el crecimiento excesivo del cerebro que se produce en ASD puede ser la base de las reducciones en la conectividad de largo alcance vistos en adolescentes y adultos con TEA ( Horwitz et al, 1988. ; Just et al, 2004. , 2007 ; Kana et al, 2007. ). El estudio actual se extiende ese trabajo a la red de análisis, que relaciona el crecimiento excesivo del cerebro en los ASD a la organización global de la red.
Las medidas de eficiencia utilizado aquí no corresponden directamente a la conectividad, la eficiencia se define en términos de caminos a través de una red, no los puntos fuertes de las conexiones individuales. Las medidas de redes capturan los aspectos más complejos de la organización del cerebro. Las ineficiencias de ASD mostrados aquí sugieren una organización en red más al azar, proporcionando el procesamiento local menos segregado y una reducción de la capacidad de integrar la información a través de la red. Las reducciones en la eficiencia global de nodal podrían provenir de cualquiera de las conexiones más débiles en general, los caminos más largos entre los nodos, o ambos. Medidas topológicas muestran más corta longitud del camino característico de ASD ( Rudie et al., 2013 ), lo que significa que la comunicación entre pares de nodos es más directo. Junto con las reducciones en la eficiencia global de nodal que se muestra aquí, esto implica una configuración más al azar, con más, pero más débiles atajos. Las reducciones en la eficiencia local, nodal apoyan esta interpretación. Dado que el grado en el que un nodo es un vecino de otro está determinado por la fuerza de la conexión directa entre ellos, los vecinos de un nodo pueden ser espacialmente distante. La eficiencia local de un nodo refleja así la agrupación espacial de sus vecinos, así como la fuerza de las conexiones entre ellos. Medidas topológicas muestran reducciones en la modularidad en TEA ( Rudie et al., 2013 ), por tanto, las reducciones en la eficiencia local de ganglionar en TEA no deben ser interpretados como de corta distancia bajo-conectividad, pero como indicativo de una configuración más al azar con un procesamiento más difusa clusters. Las ineficiencias en ASD por lo tanto sugieren a la vez menos segregación y una menor integración. La relación inversa entre las medidas de eficiencia y de ICV sugiere que estos aspectos de la organización de la red se ven afectadas por las diferencias en las trayectorias de crecimiento del cerebro.
Este estudio complementa el importante cuerpo de investigación que muestra una fuerte relación entre el tamaño del cerebro y la estructura del cerebro ( Torre, 1954 ; Jerison, 1982 ; Ringo, 1991 ; Prothero, 1997 ; Zhang y Sejnowski, 2000 ; Karbowski, 2003 ; Changizi, 2007 ; Lewis et . al, 2009 ). Que la investigación deja sin respuesta la cuestión de la etiología de estas relaciones de escala. Hemos planteado la hipótesis de que al menos algunas de estas relaciones de escala se producen sobre el desarrollo como consecuencia del impacto sobre los mecanismos normales de desarrollo de las diferencias en los costos metabólicos y retrasos en la conducción asociados con diferencias en el tamaño del cerebro ( Lewis y Elman, 2008 ; Lewis et al. , 2012 ). Nuestra hipótesis es aplicable tanto a la variabilidad individual en las trayectorias de crecimiento en el desarrollo típico, incluidas las diferencias de género, así como a las variaciones atípicas que están generalmente presentes en los trastornos del desarrollo. Los resultados actuales apoyan esta conjetura.
ICV, sin embargo, es un índice muy crudo de un fenómeno muy complejo. En los bebés con un desarrollo normal del cerebro aumenta de aproximadamente 25 por ciento de su tamaño adulto en el nacimiento hasta aproximadamente el 75 por ciento de su tamaño adulto a los 2 años de edad, con una variabilidad individual considerable en la tasa de crecimiento, así como el tamaño del cerebro maduro ( Blinkov y Glezer, 1968 ; Dobbing y Sands, 1973 ; Courchesne et al, 2000. ). Se requieren varios parámetros para capturar incluso los aspectos más básicos de estas trayectorias. ICV proporciona sólo un índice de tamaño máximo cerebro durante el desarrollo. Del mismo modo, la verdadera eficacia de la comunicación está determinada por retrasos de la conducción y los costos metabólicos, y las medidas utilizadas aquí sólo sirven como proxies de crudo para este tipo de propiedades. Las propiedades biofísicas que determinan retrasos en la conducción y los costos metabólicos, tales como la densidad de las fibras, los diámetros de los axones, y el grado de mielinización, están sólo débilmente relacionados con los resultados tractografía probabilísticos utilizados aquí como fuerzas de conexión. Además, el grado en que los resultados presentados aquí son robustos a la variedad de factores que influyen en las estimaciones basadas en tractografía de conectividad, por ejemplo, los protocolos de exploración, parámetros tractografía y parcelación de destino ( Jones et al., 2012 ), aún no se ha explorado . Las relaciones inversas entre ICV y la eficiencia por lo tanto sugieren que las trayectorias de crecimiento cerebral pueden ser responsables de una parte sustancial de las diferencias individuales en la organización del cerebro, tanto en adultos típicos, así como aquellos con TEA, pero la conjetura se deben probar además métodos que pueden proporcionar más la utilización de estimaciones precisas de las trayectorias de crecimiento cerebral, costos metabólicos, y retrasos de la conducción.
Declaración de Conflicto de Intereses
Los autores declaran que la investigación se llevó a cabo en ausencia de cualquier relación comercial o financiero que puedan interpretarse como un posible conflicto de intereses.
Agradecimientos
Esta investigación fue apoyada por subvenciones NIH / NINDS R01 NS42639, adjudicado a Jeanne Townsend. Los cálculos se realizaron sobre la supercomputadora guillimin al CLUMEQ HPC Consorcio ( http://www.clumeq.ca ). Bajo los auspicios de Compute Canadá, CLUMEQ es financiado por la Fundación Canadiense para la Innovación (CFI), el Gobierno de Québec, el Nacional de Ciencias e Ingeniería de Investigación (NSERC), y los Fonds Québécois de Recherche sur la Nature et les Tecnologías (FQRNT ).
Apraxia o dispraxia verbal: Descripción y orientaciones de intervención. Por María Gortázar Díaz
Dislexia Diagnóstico y tratamiento Escrito por el personal de Mayo Clinic
Apraxia del habla infantil Escrito por el personal de Mayo Clinic
La ansiedad en el salón de clases por Rachel Ehmke
Contener los berrinches versus manejar las crisis emocionales por Amanda Morin
Trastornos de ansiedad y TDHA por healthychildren.org American Academy of Pediatrics
Trastornos de ansiedad Escrito por el personal de Mayo Clinic
Trastorno negativista desafiante Escrito por el personal de Mayo Clinic